La fase G1 (G significa brecha) abarca desde el final de la mitosis hasta el inicio de la fase S. Durante la fase G1, las células comprueban el entorno externo y el estado intracelular, y deciden si continúan con el ciclo celular o no. En los organismos pluricelulares, el progreso del ciclo celular está influido principalmente por señales extracelulares, como la adhesión celular, o por moléculas liberadas por otras células, como los factores tróficos. También hay información interna sobre el estado de la célula que afecta al progreso del ciclo celular, como el estado de salud de la célula, si hay un número correcto de componentes celulares después de la división celular o si los cromosomas se han segregado correctamente. Cuando las señales externas e internas son correctas, las células proliferantes crecen y se preparan para entrar en la fase S.
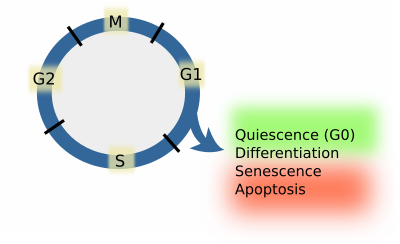
Sin embargo, las células de los organismos multicelulares no proliferan, sino que abandonan el ciclo celular a partir de la fase G1, ya sea de forma transitoria o permanente. Detener el ciclo celular significa que las células van a diferenciarse, permanecer quiescentes, pasar por procesos de senescencia o morir por apoptosis (Figura 1). Cuando las células permanecen en estado quiescente, se dice que están en fase G0. A partir de los estados G0 y diferenciado, algunos tipos de células son capaces de reiniciar el ciclo celular. La entrada en el estado quiescente supone la expresión de un conjunto particular de genes y la represión de aquellos genes que promueven la diferenciación, la senescencia o la apoptosis. Las células quiescentes reprimen los genes que promueven el ciclo celular. Desde la apoptosis o la senescencia, las células no pueden volver al ciclo celular. Por lo tanto, estas son cuatro posibles decisiones que las células pueden tomar durante la fase G1, que depende de complejos moleculares conocidos como puntos de control. Las células deben pasar estos puntos de control antes de entrar en la fase S. Si las células no pasan por uno de ellos, se dice que se ha tomado una decisión. Sin embargo, si las células no se detienen en ninguno de ellos, la fase S acabará por iniciarse. Este es el proceso por defecto. Los mecanismos moleculares en los puntos de control tienen que ser rápidos, completos e irreversibles.
Los puntos de control son complejos moleculares que controlan el progreso del ciclo celular. En el núcleo de los puntos de control se encuentran las quinasas dependientes de ciclina (CdKs). Se han encontrado nueve CdKs diferentes en los eucariotas. Para estar activas, las CdKs necesitan unirse a una proteína reguladora conocida como ciclina, y ser fosforiladas también. Una vez activadas, las CdKs fosforilan varios sustratos, como los inhibidores del ciclo celular, permitiendo así el progreso del ciclo celular. Las ciclinas son proteínas que se sintetizan periódicamente durante el ciclo celular. Existen 16 ciclinas diferentes en las células eucariotas. Las ciclinas A, B, D y E son las que más influyen en el progreso del ciclo celular. Las ciclinas D (hay 3) y E (hay 2) son importantes para el progreso de la fase G1. CdK4/ciclina D (D/CDK4) y CdK2/ciclina E (E/CDK2) fosforilan el factor de transcripción retinoblastoma, que forma parte del último punto de control de la fase G1.

El punto de control en el que se fosforila el retinoblastoma se conoce como punto de restricción porque, si se supera este punto de control, las células comienzan inmediatamente la fase S. El concepto de punto de restricción fue introducido por A. Pardee en 1974. Es muy importante porque no hay vuelta atrás, una vez iniciada la fase S, las células se dividirán y ninguna señal externa detendrá el ciclo celular. Para alcanzar el punto de restricción durante la fase G1, las células necesitan señales extracelulares como los mitógenos, pero no son necesarias una vez que se supera este punto de restricción. La secuencia es la siguiente. Los mitógenos activan receptores de la membrana plasmática que estimulan las Ras-GTPasas, que promueven la expresión de genes de factores de transcripción como c-Myc y otros. Los factores de transcripción impulsan la expresión de ciclina D que a su vez activa CdK4 y 6, que fosforilan proteínas del retinoblastoma que liberan el factor E2F (Figura 2).
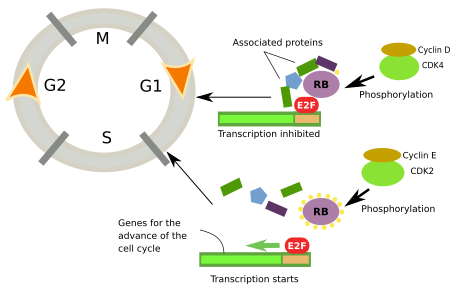
En el núcleo del punto de restricción se encuentran la CdK-ciclina, el retinoblastoma y el E2F (Figura 2). El retinoblastoma no se fosforila durante la fase G1 temprana. En esta forma, el retinoblastoma se une al E2F e inhibe la expresión de los genes que promueven el progreso del ciclo celular. El retinoblastoma es fosforilado primero por CdK4/ciclina D y luego por CdK2/ciclina E, ambos en la fase G1. El retinoblastoma tiene 16 sitios de fosforilación, lo que indica la complejidad de su regulación. La fosforilación de todos los sitios es sucesiva, y cada CdK fosforila sitios específicos. Parece que cuando se fosforilan 14 sitios, la afinidad del retinoblastoma por el E2F disminuye y, por lo tanto, el factor E2F puede promover la expresión de los genes que favorecen el inicio de la fase S.
En este entramado molecular se integran las señales que llevan información sobre el entorno (nutrientes, señales tróficas y otras), los daños en el ADN o sobre el tamaño adecuado para la división. Si todo es correcto, la célula pasa el punto de restricción y comienza la fase S. Si algo va mal, hay varios tipos de inhibidores que detienen el avance del ciclo celular. Uno de ellos es el p53, que se activa en muchos tumores. Cuando el ADN está dañado, o la célula está sometida a estrés, o hay cambios en el pH, o se da cualquier otra situación potencialmente peligrosa para la célula, p53 se sobreexpresa y activa el gen p21, que a su vez inhibe la fosforilación del retinoblastoma, por lo que el ciclo celular se detiene y no comienza la fase S.
Como hemos mencionado anteriormente, la mayoría de las células de un organismo pluricelular no están en permanente proliferación. Esto se debe a que existen inhibidores de Cdk/ciclinas en fase G1 que hacen que las células abandonen el ciclo celular.
Bibliografía
Blomen VA, Boonstra J. 2007. Determinación del destino celular durante la progresión de la fase G1. Cellular and molecular life sciences. 64:3084-3104.
Matson JP, Cook JG. 2017. Decisiones de proliferación del ciclo celular: el impacto de los análisis de células individuales. The FEBS journal. 284: 362-375.
0 comentarios