Acabou de ler sobre duas vias no catabolismo da glicose – a glicólise e o ciclo do ácido cítrico – que geram o ATP. A maior parte do ATP gerado durante o catabolismo aeróbico da glucose, no entanto, não é gerado directamente a partir destas vias. Em vez disso, deriva de um processo que começa com a passagem de electrões através de uma série de reacções químicas a um aceitador final de electrões, o oxigénio. Este é o único local na respiração aeróbica onde o O2 é realmente necessário. Estas reacções ocorrem em complexos proteicos especializados localizados na membrana interna das mitocôndrias dos organismos eucarióticos e na parte interna da membrana celular dos organismos procarióticos. A energia dos electrões é utilizada para gerar ATP. A totalidade deste processo chama-se fosforilação oxidativa.
Fosforilação oxidativa duradoura:
- A energia de NADH e FADH2 é utilizada até.
- O oxigénio gas é convertido em água.
- 30-36 ATP são recarregados de ADP
A cadeia de transporte de electrões (Figura 1) é o último componente da respiração aeróbica e é a única parte do metabolismo que utiliza o oxigénio atmosférico. O oxigénio difunde-se continuamente nas plantas para este fim. Nos animais, o oxigénio entra no corpo através do sistema respiratório. O transporte de electrões é uma série de reacções químicas que se assemelham a uma brigada de baldes em que os electrões passam rapidamente de um componente para o outro, para o ponto final da cadeia, onde o oxigénio é o último aceiro de electrões e a água é produzida. Existem quatro complexos compostos de proteínas, rotulados I a IV na Figura 1, e a agregação destes quatro complexos, juntamente com portadores de electrões móveis e acessórios associados, é chamada cadeia de transporte de electrões. A cadeia de transporte de electrões está presente em múltiplas cópias na membrana mitocondrial interna de eucariotas e na membrana plasmática de procariotas. Em cada transferência de um electrão através da cadeia de transporte de electrões, o electrão perde energia, mas com algumas transferências, a energia é armazenada como energia potencial, utilizando-a para bombear iões de hidrogénio (H+, prótons) através da membrana mitocondrial interna para o espaço intermembrana, criando um gradiente electroquímico. Um gradiente electroquímico consiste em duas partes: uma diferença na concentração de soluto através da membrana combinada com uma diferença na carga através da membrana. Aqui, o gradiente electroquímico é constituído por uma maior concentração de H+ no espaço interior da membrana, em comparação com a matriz mitocondrial.
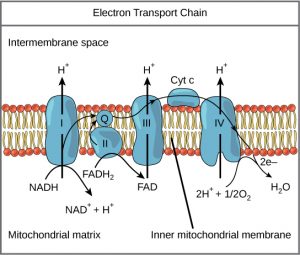
Electrões de NADH e FADH2 são passados para complexos proteicos na cadeia de transporte de electrões. Como são passados de um complexo para outro (há um total de quatro), os electrões perdem energia, e alguma dessa energia é utilizada para bombear iões de hidrogénio da matriz mitocondrial para o espaço intermembrana. No quarto complexo proteico, os electrões são aceites pelo oxigénio, o aceitador terminal. O oxigénio com os seus electrões extra combina-se então com dois iões de hidrogénio, aumentando ainda mais o gradiente electroquímico, para formar água. Se não houvesse oxigénio presente nas mitocôndrias, os electrões não poderiam ser removidos do sistema, e toda a cadeia de transporte de electrões faria marcha atrás e pararia. As mitocôndrias seriam incapazes de gerar novos ATP desta forma, e a célula acabaria por morrer por falta de energia. Esta é a razão pela qual temos de respirar para extrair novo oxigénio. Este é o único local onde o oxigénio é necessário durante os processos de respiração aeróbica.
Na cadeia de transporte de electrões, a energia livre da série de reacções que acabamos de descrever é utilizada para bombear iões de hidrogénio através da membrana. A distribuição desigual dos iões H+ através da membrana estabelece um gradiente electroquímico, devido à carga positiva dos iões H+ e à sua maior concentração num dos lados da membrana.
Iões de hidrogénio difundem-se do espaço intermembrana através da membrana interna para a matriz mitocondrial através de uma proteína de membrana integral chamada ATP synthase (Figura 2). Esta proteína complexa actua como um minúsculo gerador, virado pela força dos iões de hidrogénio que se difundem através dela, descendo o seu gradiente electroquímico a partir do espaço da membrana intermembrana, onde há muitos iões de hidrogénio que se repelem mutuamente para a matriz, onde são poucos. O giro das partes desta máquina molecular regenera o ATP a partir do ADP. Este fluxo de iões de hidrogénio através da membrana através da ATP synthase é chamado quimiossimose.
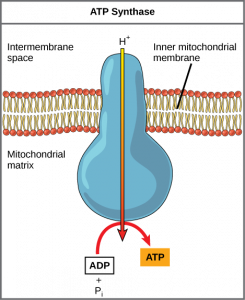
Chemiosmosmosis (Figura 2) é utilizada para gerar 90 por cento do ATP feito durante o catabolismo aeróbico da glucose. O resultado das reacções é a produção de ATP a partir da energia dos electrões removidos dos átomos de hidrogénio. Estes átomos eram originalmente parte de uma molécula de glucose. No final do sistema de transporte de electrões, os electrões são utilizados para reduzir uma molécula de oxigénio a iões de oxigénio. Os electrões extra nos iões de oxigénio atraem iões de hidrogénio (prótons) do meio circundante, e a água é formada. A cadeia de transporte de electrões e a produção de ATP através da quimiossimose são colectivamente chamadas fosforilação oxidativa (Figura 3).
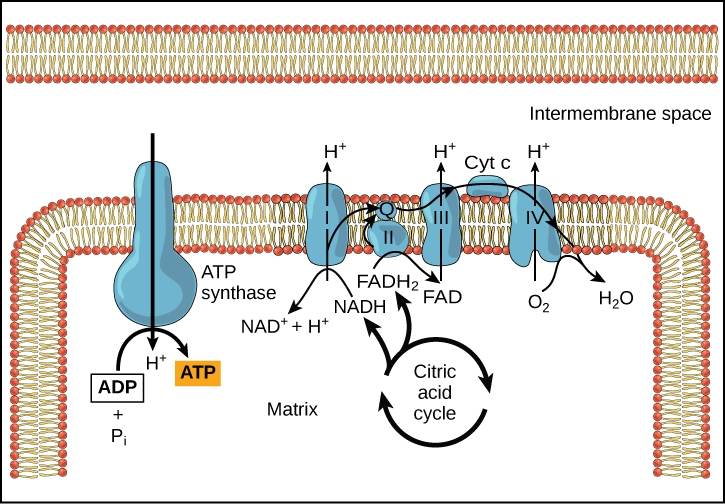
h2>ATP Yield
O número de moléculas ATP geradas a partir do catabolismo da glucose varia. Por exemplo, o número de iões de hidrogénio que os complexos de cadeias de transporte de electrões podem bombear através da membrana varia de espécie para espécie. Outra fonte de variação deriva do vaivém de electrões através das membranas das mitocôndrias porque o NADH gerado a partir da glicólise não pode entrar facilmente nas mitocôndrias. Assim, os electrões são captados no interior das mitocôndrias ou por NAD+ ou FAD+. Como já foi dito anteriormente, estas FAD+molecules podem transportar menos iões; consequentemente, são geradas menos moléculas de ATP quando o FAD+ actua como um transportador. O NAD+ é utilizado como transportador de electrões no fígado e o FAD+ actua no cérebro.
Outro factor que afecta o rendimento das moléculas de ATP geradas a partir da glucose é o facto de os compostos intermediários nestas vias serem utilizados para outros fins. O catabolismo da glucose liga-se às vias que constroem ou decompõem todos os outros compostos bioquímicos nas células, e o resultado é um pouco mais confuso do que as situações ideais descritas até agora. Por exemplo, os açúcares que não a glicose são introduzidos na via glicolítica para a extracção de energia. Além disso, os açúcares de cinco carbonos que formam ácidos nucleicos são feitos de intermediários na glicólise. Certos aminoácidos não essenciais podem ser feitos a partir de intermediários tanto da glicólise como do ciclo do ácido cítrico. Lípidos, tais como colesterol e triglicéridos, são também feitos a partir de intermediários nestas vias, e tanto os aminoácidos como os triglicéridos são decompostos para energia através destas vias. Em geral, nos sistemas vivos, estas vias do catabolismo da glucose extraem cerca de 34% da energia contida na glucose.
Secção Resumo
A cadeia de transporte dos electrões é a porção da respiração aeróbica que utiliza oxigénio livre como o aceitador final dos electrões removidos dos compostos intermediários do catabolismo da glucose. A cadeia de transporte de electrões é composta por quatro grandes complexos multiproteicos embutidos na membrana mitocondrial interna e dois pequenos portadores difusíveis de electrões que fecham os electrões entre eles. Os electrões são passados através de uma série de reacções, com uma pequena quantidade de energia livre utilizada em três pontos para transportar iões de hidrogénio através de uma membrana. Este processo contribui para o gradiente utilizado na quimiossimose. Os electrões que passam através da cadeia de transporte de electrões perdem gradualmente energia até acabarem por ser doados ao oxigénio gasoso que aceita dois protões (H+) e é convertido em água. Os produtos finais da cadeia de transporte de electrões são água e cerca de 30-34 moléculas de ATP. Vários compostos intermediários do ciclo do ácido cítrico podem ser desviados para o anabolismo de outras moléculas bioquímicas, tais como aminoácidos não essenciais, açúcares, e lípidos. Estas mesmas moléculas podem servir como fontes de energia para as vias da glucose.
0 comentários