La phase G1 (G signifie gap) s’étend de la fin de la mitose au début de la phase S. Pendant la phase G1, les cellules vérifient l’environnement extérieur et l’état intracellulaire, et décident si elles poursuivent ou non le cycle cellulaire. Dans les organismes multicellulaires, la progression du cycle cellulaire est principalement influencée par des signaux extracellulaires, comme l’adhésion cellulaire, ou des molécules libérées par d’autres cellules, comme les facteurs trophiques. Il existe également des informations internes sur l’état de la cellule qui affectent la progression du cycle cellulaire, comme l’état de santé de la cellule, le nombre correct de composants cellulaires après la division cellulaire ou la ségrégation correcte des chromosomes. Lorsque les signaux externes et internes sont bons, les cellules en prolifération se développent et se préparent à entrer en phase S.
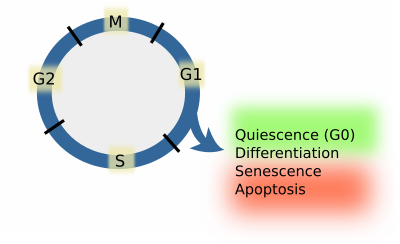
Cependant, les cellules des organismes multicellulaires ne prolifèrent pas, mais quittent le cycle cellulaire à partir de la phase G1, de façon transitoire ou permanente. L’arrêt du cycle cellulaire signifie que les cellules vont se différencier, rester quiescentes, passer par des processus de sénescence ou mourir par apoptose (figure 1). Lorsque les cellules restent dans un état quiescent, on dit qu’elles sont en phase G0. À partir des états G0 et différencié, certains types de cellules sont capables de redémarrer le cycle cellulaire. L’entrée en phase quiescente signifie l’expression d’un ensemble particulier de gènes et la répression des gènes qui favorisent la différenciation, la sénescence ou l’apoptose. Les cellules quiescentes répriment les gènes qui favorisent le cycle cellulaire. À partir de l’apoptose ou de la sénescence, les cellules ne peuvent plus retourner dans le cycle cellulaire. Il s’agit donc de quatre décisions possibles que les cellules peuvent prendre pendant la phase G1, qui dépend de complexes moléculaires appelés points de contrôle. Les cellules doivent passer ces points de contrôle avant d’entrer dans la phase S. Si les cellules ne passent pas l’un d’entre eux, on dit qu’une décision a été prise. En revanche, si les cellules ne s’arrêtent à aucun d’entre eux, la phase S finira par démarrer. Il s’agit du processus par défaut. Les mécanismes moléculaires aux points de contrôle doivent être rapides, complets et irréversibles.
Les points de contrôle sont des complexes moléculaires qui contrôlent la progression du cycle cellulaire. Au cœur des points de contrôle se trouvent les kinases cyclines-dépendantes (CdKs). Neuf CdKs différentes ont été trouvées chez les eucaryotes. Pour être actives, les CdK doivent se lier à une protéine régulatrice appelée cycline, et être également phosphorylées. Une fois activées, les CdK phosphorylent plusieurs substrats, tels que les inhibiteurs du cycle cellulaire, permettant ainsi la progression du cycle cellulaire. Les cyclines sont des protéines synthétisées périodiquement au cours du cycle cellulaire. Il existe 16 cyclines différentes dans les cellules eucaryotes. Les cyclines A, B, D et E ont le plus d’impact sur la progression du cycle cellulaire. Les cyclines D (il y en a 3) et E (il y en a 2) sont importantes pour la progression de la phase G1. La CdK4/cycline D (D/CDK4) et la CdK2/cycline E (E/CDK2) phosphorent le facteur de transcription rétinoblastome, qui fait partie du dernier point de contrôle de la phase G1.

Le point de contrôle où le rétinoblastome est phosphorylé est connu sous le nom de point de restriction car, si ce point de contrôle est passé, les cellules commencent immédiatement la phase S. Le concept de point de restriction a été introduit par A. Pardee en 1974. Il est très important car il n’y a pas de retour en arrière possible, une fois que la phase S a commencé, les cellules se divisent et aucun signal externe ne peut arrêter le cycle cellulaire. Pour atteindre le point de restriction pendant la phase G1, les cellules ont besoin de signaux extracellulaires comme les mitogènes, mais ils ne sont plus nécessaires une fois que ce point de restriction est passé. La séquence est la suivante. Les mitogènes activent les récepteurs de la membrane plasmique qui stimulent les Ras-GTPases, lesquelles favorisent l’expression des gènes des facteurs de transcription comme c-Myc et autres. Les facteurs de transcription stimulent l’expression de la cycline D qui, à son tour, active les CdK4 et 6, lesquelles phosphorylent les protéines du rétinoblastome qui libèrent le facteur E2F (figure 2).
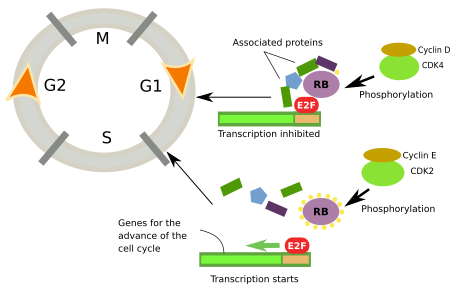
Au cœur du point de restriction se trouvent la CdK-cycline, le rétinoblastome et E2F (figure 2). Le rétinoblastome n’est pas phosphorylé au début de la phase G1. Sous cette forme, le rétinoblastome est lié à E2F et inhibe l’expression des gènes qui favorisent la progression du cycle cellulaire. Le rétinoblastome est d’abord phosphorylé par CdK4/cycline D, puis par CdK2/cycline E, tous deux en phase G1. Le rétinoblastome possède 16 sites de phosphorylation, ce qui indique la complexité de la régulation. La phosphorylation de tous les sites est successive, et chaque CdK phosphoryle des sites spécifiques. Il semble que lorsque 14 sites sont phosphorylés, l’affinité du rétinoblastome pour E2F diminue et, par conséquent, le facteur E2F peut favoriser l’expression de gènes qui favorisent le début de la phase S.
Dans ce cadre moléculaire, sont intégrés les signaux porteurs d’informations sur l’environnement (nutriments, signaux trophiques et autres), sur les dommages à l’ADN ou sur la taille adéquate pour la division. Si tout est correct, la cellule passe le point de restriction et la phase S commence. Si quelque chose ne va pas, il existe plusieurs types d’inhibiteurs qui arrêtent la progression du cycle cellulaire. L’un d’eux est le p53, qui est activé dans de nombreuses tumeurs. Lorsque l’ADN est endommagé, ou que la cellule est soumise à un stress, ou qu’il y a des changements dans le pH, ou toute autre situation potentiellement dangereuse pour la cellule sont présents, p53 est surexprimé et active le gène p21, qui à son tour inhibe la phosphorylation du rétinoblastome, de sorte que le cycle cellulaire est arrêté et ne commence pas la phase S.
Comme nous l’avons mentionné plus haut, la plupart des cellules d’un organisme multicellulaire ne sont pas en prolifération permanente. En effet, il existe des inhibiteurs de Cdk/cyclines en phase G1 qui font que les cellules quittent le cycle cellulaire.
Bibliographie
Blomen VA, Boonstra J. 2007. Détermination du destin cellulaire pendant la progression de la phase G1. Sciences de la vie cellulaires et moléculaires. 64:3084-3104.
Matson JP, Cook JG. 2017. Décisions de prolifération du cycle cellulaire : l’impact des analyses de cellules uniques. The FEBS journal. 284 : 362-375.
0 commentaire